OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
TRANSPORTNI MEHANIZMI ĆELIJSKE MEMBRANE
(AKTIVNI I PASIVNI TRANSPORT)
Ćelijska citoplazma je od okoline odvojena polupropustljivom ćelijskom
membranom. Ćelijska membrana se sastoji od dvostrukog lipidnog sloja u
koji su uronjeni različiti proteini. Lipidni deo se sastoji od fosfolipida,
holesterola i glikolipida. Fosfolipidi ćelijske membrane sastoje se iz
hidrofilne (polarne) glave i hidrofobnog (nepolarnog) repa, te zahvaljujući
tome spontano formiraju dva sloja u ćelijskoj membrani tako što se hidrofobni
repovi grupišu u unutrašnjosti, a polarne glave orjentišu se prema spoljašnjoj
strani ćelijske membrane. (1)
Proteini uronjeni u lipidni sloj mogu se podjeliti
u dve vrste:
- Integralni proteini su transmembranski proteini, koji imaju ulogu:
- jonskih kanala kroz koje difunduju mali hidrofilni molekuli i joni,
- transportnih nosača u pasivnom i aktivnom transportu,
- pumpi u aktivnom transportu jona nasuprot elektrohemijskog gradijenta,
- receptora u unutarćelijskim reakcijama;
- Periferni proteini na površini lipidnog dvosloja (nalaze se samo sa jedne strane membrane), čija je uloga uglavnom receptorna (za hormone, antigene, neurotransmitere). (2)
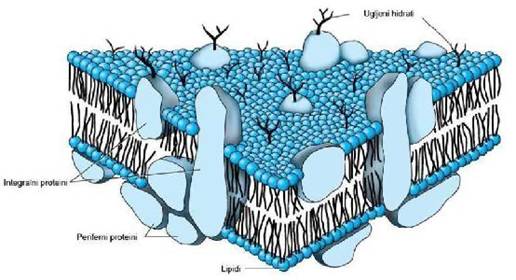
Slika 1.1. Struktura ćelijske membrane
Molekuli kroz ćelijsku membranu prolaze na različite načine:
- prostom difuzijom prolaze gasovi (kiseonik, ugljen – dioksid i azot) i male relativno hidrofobne molekule (etanol), u skladu sa gradijentom koncentracije, tj. sa mesta više na mesto niže koncentracije. Difuzija vode kroz polupropustljivu membranu sa mesta više na mesto niže koncentracije naziva se osmoza.
- olakšanom difuzijom (pasivni transport), uz pomoć proteina tj. jonskih kanala, prolaze molekuli sa nabojem (hloridi i bikarbonati) u skladu sa gradijentom koncentracije i električnim gradijentom tj. elektrohemijskim gradijentom;
- olakšanom difuzijom (pasivni transport), uz pomoć transportnih proteina, prolaze nešto veće molekule (glukoza) u skladu sa gradijentom koncentracije;
- aktivnim transportom, uz pomoć transportnih proteina ili jonskih kanala, prolaze polarne molekule i molekule sa nabojem (pumpa za Na+ i K+) suprotno elektrohemijskom gradijentu uz utrošak energije (ATP). U zavisnosti od toga da li se za aktivni transport energija osigurava direktno ili indirektno, razlikujemo primarni i sekundarni transport. (1)
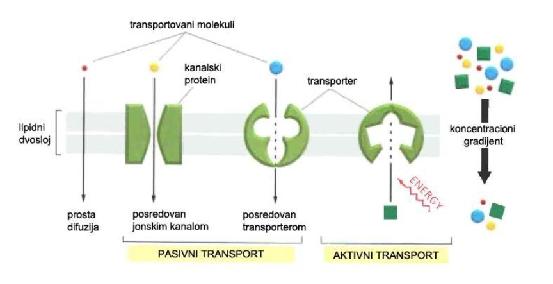
Slika 1.2. Tipovi transporta kroz membranu
Telesne tečnosti dele se na:
- intercelularna tečnost (ICT) – bogata je K+;
- ektracelularna tečnost (ECT) - bogata je Na+, Ca++ i Cl-, a čine je intersticijumska tečnost (IST) i intravaskularna tečnost (IVT) ili krvna plazma. (2)
Pasivni transport
Pasivni transport ili olakšana difuzija je proces prenosa (kroz ćelijsku membranu):
- polarnih molekula, posredstvom transportnih proteina, u skladu sa gradijentom koncentracije tj. sa mjesta više na mjesto niže koncentracije, i
- jona, posredstvom proteina tj. jonskih kanala, u skladu sa gradijentom koncentracije i elektrohemijskim gradijentom; (1)
Pasivni transport odvija se iz sredine gde je koncentracija molekula
viša u sredinu gdje je njegova koncentracija niža. Ukoliko molekul nosi
naelektrisanje, pored koncentracionog gradijenta, na njega će djelovati
i razlika u električnom potencijalu sa spoljašnje i unutrašnje strane
membrane tj. elektrohemijski gradijent. Pasivni transport nikad ne zahteva
utrošak energije.
Na primer, molekul glukoze je krupan i polaran, pa ne može difundovati
ni kroz lipidni dvosloj ni kroz pore ćelijske membrane. Glukoza je esencijalan
molekul bez kojeg ćelija ne može obavljati svoje funkcije. Kako se unutar
ćelije glukoza troši u metaboličkim procesima, njena koncentracija u interćelijskoj
tečnosti je manja nego u ekstracelularnoj tečnosti. Glukoza se transportuje
pomoću transportnog proteina glukoze (Glut tj. facilitirajuće transportere
glukoze), za čiji kanal se vezuje na receptornom mestu, transportni protein
pri tome doživljava konformacijsku promenu koja se završava otvaranjem
kanala s unutrašnje strane membrane. (2)
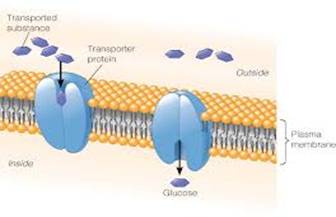
Slika 2.1.1. Olakšana difuzija glukoze kroz ćelijsku membranu
Procesom olakšane difuzije, usljed elektrohemijskog gradijenta kroz ćelijsku membranu prolaze hloridi i bikarbonati, što je važno za održavanje acido-bazne ravnoteže u organizmu. (2)
Aktivni transport
Molekuli se u aktivnom transportu prenose posredstvom proteina nasuprot elektrohemijskog gradijenta pa je za aktivni transport neophodan utrošak energije što podrazumjeva direktan utrošak ATP-a (primarni aktivni transport), ili najčešće kotransport jona kalijuma u skladu s njegovim elektrohemijskim gradijentom (koristi se enrgija jonskog gradijenta) što podrazumjeva indirektan utrošak ATP-a u pumpi za Na+ i K+ (sekundarni aktivni transport). (1)
Primarni aktivni transport
Kako proteinski transporter prenosi molekul aktivnim transportom suprotno
elktrohemijskom gradijentu, mora utrošiti energiju. Energiju troši direktnom
hidrolizom ATP-a na ADP i P, što znači da ima ATP-aznu aktivnost (kao
enzim). Pošto je ATP direktno povezan sa procesom transporta ovakav aktivni
transport se naziva primarnim. (2)
Na primjer, koncentracija jona natrijuma je veća u ekstracelularnoj tečnosti
u odnosu na intracelularnu tečnost, pa joni natrijuma imaju tendenciju
da ulaze u ćeliju olakšanom difuzijom. Koncentracija jona kalijuma je
veća u intracelularnoj tečnosti u odnosu na ekstracelularnu tečnost, pa
joni kalijuma imaju tendenciju da izađu iz ćelije olakšanom difuzijom.
Pumpa za Na+ i K+ održava odgovarajuće koncentracije jona natrijuma i
jona kalijuma u ćeliji, te ispumpava 3 jona natrijuma iz ćelije (dakle
nasuprot gradijentu koncentracije jona natrijuma) i upumpava u ćeliju
2 jona kalijuma ( također nasuprot gradijentu koncentracije). Ovaj proces
je moguć samo uz utrošak energije tj. hidroze ATP-a na proteinskom kanalu
za transport natrijuma i kalijuma, koji pri tom mjenja svoju konformaciju.
Pošto se energija troši na samom proteinskom nosaču, ovaj je proces primjer
primarnog aktivnog transporta. (2)
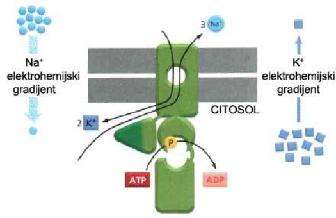
Slika 2.2.1.1. Pumpa za natrijum i kalijum
Pored pumpe za Na+ i K+, primjer primarnog aktivnog transporta je i pumpa za Ca++, koja je karakteristična za membrane sarkoplazmatskog retikuluma, čija je unutrašnjost bogata jonima kalcijuma, dok je citoplazma siromašna jonima kalcijuma. (2)
Sekundarni aktivni transport
Sekundarni aktivni transport nekog molekula odvija se posredstvom proteina transportera suprotno elektrohemijskom gradijentu, pri čemu se troši energija indirektno tj. troši se energija elektrohemijskog gradijenta drugog jona. Transport molekula koji se prenosi suprotno svom elektrohemijskom gradijentu, zavisi od transporta drugog molekula koji se prenosi u smeru elektrohemijskog gradijenta. Proteinski transporter ima dva vezna mesta, a do njegove konformacijske promene dolazi tek nakon vezivanja oba molekula, čime se isporučuju oba molekula. U zavisnosti od smera kojim se oba molekula transportuju razlikuje se:
- Kotransport (simport) kada se oba molekula transportuju u istom smeru,
- Kontratransport (antiport) kada se molekuli transportuju u različitim smerovima. (2)
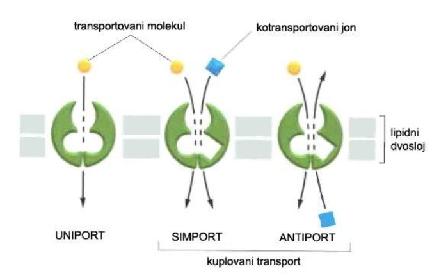
Slika 2.2.2.1. Tipovi aktivnog transporta
Primer sekundarnog aktivnog kotransporta (simporta) je natrijum
- zavisan proteinski transporter za glukozu (za vreme varenja na luminalnoj
membrani). Na početku intercelularna tečnost je siromašna glukozom, pa
glukoza u ćeliju ulazi olakšanom difuzijom. Međutim, kada se dostigne
difuziona ravnoteža s obe strane membrane, aktivira se sekundarni aktivni
transport. Glukoza se u ovom slučaju kreće suprotno svom gradijentu koncentracije,
jer je intercelularna tečnost bogata glukozom. Natrijumov jon kretat će
se u smeru svog elektrohemijskog gradijenta. Molekul glukoze (transportovani
molekul) i natrijumov jon (kotransportovani jon) transportuju se u istom
smeru. Naknadno će pumpa za Na+ i K+ uz utrošak energije (hidrolizom ATP-a)
iz intracelularne tečnosti ispumpati višak jona natrijuma. (2)
Primjer sekundarnog aktivnog kontratransporta (antiporta) je protein transporter
za natrijumov i kalcijumov jon na ćelijskoj membrani ćelija srčanog mišića.
Natrijumov jon vezuje se za proteinski transporter sa spoljašnje strane
membrane, a kalcijumov jon sa unutrašnje strane membrane. Kalcijumov jon
se iz intracelularne tečnosti transportuje nasuprot elektrohemijskom gradijentu,
a natrijumov jon u skladu sa elektrohemijskim gradijentom u intracelurnu
tečnost. Pumpa za Na+ i K+ će uz utrošak energije (hidrolizom ATP-a) iz
intracelularne tečnosti ispumpati višak jona natrijuma. (2)
Ćelijska membrana svojom selektivnom propustljivošću
i procesima pasivnog i aktivnog transporta, od ključnog je značaja za
održavanje transmembranskog gradijenta koncentracije niza molekula i jona,
a time i za održavanje i regulaciju normalnog života ćelije, kao i organizma
u celini. (2)
Koncentracioni gradijent jona kalijuma kroz ćelijsku membranu neophodan
je za uspostavljanje mirovnog membranskog potencijala nerva i mišića.
Visoka koncentracija jona kalijuma u intercelularnoj tečnosti neophodna
je za rast i deobu ćelije i funkciju mnogih enzima. Koncentracioni gradijent
jona natrijuma kroz ćelijsku membranu neophodan je za uspostavljanje akcionog
potencijala. Apsorpcija glukoze i aminokiselina zavisi od gradijenta koncentracije
kroz ćelijsku membranu natrijuma. Ekscitacija i kontrakcija mišićne ćelije
zavisi od razlike u koncentraciji jona kalcijuma kroz membrane sarkoplazmatičnog
retikuluma. Transmembranski elektrohemijski gradijent hlorida i bikarbonata
važan je za održavanje acido-bazne ravnoteže organizma. (2)
Stalni rad pumpe za Na+ i K+ doprinosi normalnom volumenu ćelije, kao
i normalnom transmembranskom koncentracionom gradijentu jona kalijuma
i natrijuma, te bi život ćelije bez pumpe za Na+ i K+ bio nemoguć. (2)
- Diklić, V., Kosanović, M., Nikoliš, J., Dukić, S.: Biologija sa humanom genetikom – reprint izd. Iz 1997, - Beograd: Grafopan, 2001.
- Mujović, V.: Medicinska fiziologija – Beograd: A-Z BOOK, 2004
Student: Dragana Martinović
preuzmi
seminarski rad u wordu » » »

Besplatni Seminarski
Radovi