OSTALI SEMINARSKI RADOVI
IZ VETERINE: |
|||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||
OKSIDATIVNI STRES MLIJEČNIH KRAVA U
PERIPARTALNOM PERIODU
Oksidativni stres se definiše kao poremećaj u odnosu između oksidanasa i antioksidanasa, bilo na nivou pojedinačne ćelije ili cijelog organizma (Lykkesfeldt, 2007). Prema Siesu (1991) oksidativni stres nastaje kao posljedica disbalansa između proizvodnje reaktivnih metabolita kiseonika (Reactive Oxygen Metabolites, ROM ili Reactive Oxygen Species, ROS) i kapaciteta antioksidativnih mehanizama za njihovu neutralizaciju. Kao rezultat tog disbalansa dolazi do oksidativnog oštećenja ćelija i tkiva koje se može manifestovati oksidativnom modifikacijom ćelijskih makromolekula, programiranom ćelijskom smrću – apoptozom i strukturnim oštećenjem tkiva. Toyokuni (1999) navodi da oksidativni stres izaziva peroksidativno oštećenje lipida i ostalih ćelijskih makromolekula, što ima za posljedicu oštećenje ćelijske membrane i drugih dijelova ćelije. Iako postoji više podjela oksidanasa, od kojih se najčešće pominje ona na radikale i ne-radikale (hemijska podjela), za kliničku praksu je najprimjenjljivija ona koju je dao Buettner (1993). Ovaj autor je sve oksidanse podijelio na osnovu njihove sposobnosti da odvoje elektron od supstrata, tj. prema njihovoj potentnosti i biološkoj reaktivnosti, a time i potencijalnoj škodljivosti za organizam. Prema ovoj klasifikaciji, kao najpotentniji oksidans označen je hidroksilni jon.
Tokom fizioloških procesa oslobađa se čitav niz oksidišućih supstanci, pri čemu je njihova količina u granicama koje, u kombinaciji sa aktivacijom antioksidativne odbrane, ne dovode do oštećenja ćelije i organizma. Međutim, produkcija oksidanasa može biti povećana kao rezultat niza patoloških i ne-patoloških okolnosti, i tada dolazi do oštećenja izazvanih dejstvom tih radikala (Roth, 2000). Ćelije aerobnih organizama neprekidno su izložene i djelovanju niza egzogenih oksidansa (Haliwel and Gutterridge, 1999). Iako se pojam oksidansa i oksidativne aktivnosti redovno spominje kao negativna pojava i uzrok oštećenja ćelije treba imati na umu i njihove fiziološke uloge kao signalnih molekula uključenih u kontrolu kaskadnih procesa, kao što su apoptoza i zapaljenske reakcije.
Dakle, prisustvo viška oksidansa nije nužno povezano sa oštećenjem ćelija. Dobar primjer za to jeste NO* radikal, koji učestvuje u kontroli mikrocirkulacije (kako u fiziološkim, tako i u patološkim uslovima, odnosno tokom razvoja zapaljenja). Ipak, povišena proizvodnja ovog radikala može rezultovati povišenjem nivoa peroksinitritnog radikala koji ima štetne efekte. Ovaj primjer ukazuje da se, prilikom dokazivanja prisustva i procjene intenziteta oksidativnog stresa, zaključak ne može bazirati isključivo na vrijednostima markera oksidativnog stresa, kao što su ovi radikali, bez registrovanja nastalih oštećenja.
Glavni endogeni izvor oksidansa pod fiziološkim uslovima je respiratorni lanac elektrona u mitohondrijama iz koga stalno "curi" manja količina kiseonika u obliku ROS međuproizvoda. Ovaj izvor oksidacije formira njihov bazalni nivo u organizmu, koji se pod određenim uslovima mijenja. Jedan od faktora koji redovno podiže nivo oksidanasa u organizmu jesu imune reakcije tokom infekcije ili inflamacije i eventualnih autoimunih procesa. Ovaj izvor oksidanasa u osnovi ima oslobađanje oksidativnih supstanci iz imunoreaktivnih ćelija, prije svega neutrofila (u okviru "oksidativnog praska"), sa ciljem da se unište patogeni mikroorganizmi i drugi antigeni, potencijalno štetni za organizam. Zbog nedostatka specifičnosti i selektivnosti, ove supstance djeluju na ciljni molekul ili ćeliju, ali i na okolno tkivo, dovodeći do njegovog oštećenja. Enzimi kao što je ksantin oksidaza ili azot-oksid sintetaza proizvode (između ostalog) superoksidni radikal i NO*, čijom reakcijom može nastati posebno opasan peroksinitrit. Iako se u in vitro uslovima oksidansi mogu dobiti i iz Fentonove reakcije (oksidacija katalizovana prisustvom jona prelaznih metala), u in vivo uslovima ovo je malo vjerovatno, pa se stoga ova reakcija ne smatra značajnijim potencijalnim izvorom oksidansa u živom organizmu (Chen i sar. 2000).
Pored endogenih, veoma važan izvor oksidanasa koji mogu dovesti do nastanka oksidativnog stresa su i egzogeni izvori (jonizuće i nejonizuće zračenje, toksični gasovi i hemikalije, oksidaciona dezinfekciona sredstva, ali svakako i hrana bogata sastojcima koji lako oksidišu, kao što su polinezasićene masne kiseline).
Ciljni molekuli za dejstvo oksidanasa su proteini,
masti i DNK.
Pod dejstvom oksidativnih supstanci nastaje niz oštećenja na DNK, koja
se manifestuju prekidima jednog ili oba lanca, ugradnjom pogrešnih baza,
mutacijama i drugim promjenama (Poulsen, 2005). Oksidacija proteina
ima za posljedicu stvaranje defektnih proteina, što rezultuje poremećajem
njihovih funkcija, naročito ukoliko se radi o transportnim proteinima
ili enzimima. Masti, naročito one koje sadrže nezasićene masne kiseline,
posebno su osjetljive na oksidaciju. Unutar istog molekula i između više
molekula masti unutar npr. ćelijske membrane, pod uticajem oksidanasa
brzo i lako nastaju reakcije oksidacije, što može narušiti važne funkcije,
pa i sam integritet ćelije.
Mehanizmi odbrane od oksidativnog stresa
S obzirom na sredinu u kojoj žive, aerobni organizmi se stalno susreću sa oksidativnim supstancama, te su tokom evolucije razvili čitav niz mehanizama za odbranu od oksidativnog stresa. Generalno gledano, odbrana od oksidativnog stresa se odvija na više nivoa, kroz djelovanje antioksidativnih supstanci, popravak ili uklanjanje oštećenja nastalih dejstvom oksidanasa i inkapsulaciju ireparabilnih oštećenja, uključujući tu i apoptozu - programiranu ćelijsku smrt.
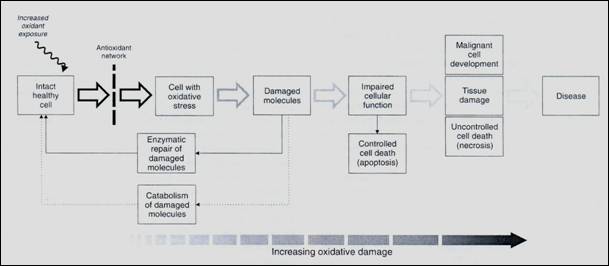
Slika 1. Šematski prikaz ćelijskih mehanizama odbrane od oksidativnog stresa (prema Lykkesfeldtu i Svendsenu, 2007)
Prvi nivo odbrane od oksidativnih supstanci jeste tzv. antioksidativna
mreža, sastavljena od niza antioksidansa, supstanci koje mogu donirati
elektron oksidansima, dovodeći njih u stabilno stanje, pri čemu se sami
oksiduju. Nastali radikali antioksidansa su mnogo manje reaktivni i ne
dovode do oštećenja bioloških molekula. Takođe, unutar antioksidativne
mreže postoje sistemi kojima se oksidisani antioksidansi recikliraju u
prvobitno stanje nizom reakcija koje koriste energiju iz NADPH. Ovo recikliranje
i reutilizacija antioksidansa je i ključ efekta antioksidativne mreže,
jer bi se bez toga njen kapacitet brzo zasitio (Lykkesfeldt i sar.
2003). Antioksidativna mreža se sastoji od dvije grupe supstanci,
antioksidativnih supstanci male i velike molekulske mase. U grupu antioksidansa
male molekulske mase spadaju glutation (GSH), vitamin E (α-tokoferol)
i vitamin C (askorbinska kiselina). Njihova aktivnost omogućena je enzimima
dehidroaskorbat reduktazom i glutation reduktazom, koje ih nakon reakcije
sa oksidansima vraćaju u neoksidisano stanje. Antioksidansi velike molekulske
mase su manje brojni i uglavnom predstavljaju enzimske sisteme, kao što
su supreoksid-dismutaza (koja katalizuje spajanje dva molekula superoksidnog
radikala u jedan molekul vodonik-peroksida i jedan molekul kiseonika),
katalaza i glutation peroksidaza (koje katalizuju razlaganje vodonik-peroksida,
koji stalno "curi" iz transportnog lanca elektrona).
Ukoliko je oštećenje već nastalo ili antioksidativna mreža nije u mogućnosti
da zaštiti ćeliju od njegovog nastanka, aktiviraju se mehanizmi koji otkrivaju
i popravljaju oštećena mjesta, ili isijecaju oštećene segmente koji zatim
bivaju podvrgnuti kataboličkim procesima. Ovi mehanizmi (uglavnom enzimske
prirode) posebno su dobro razvijeni kada je u pitanju detekcija i otklanjanje
oštećenja na DNK. Slični mehanizmi postoje i za druge sastojke ćelije,
posebni lipide ćelijske membrane, koji imaju zadatak da očuvaju integritet
ćelije.
U slučaju da su oštećenja prevazišla kapacitet ćelije za popravku ili
otklanjanje oštećenja putem isijecanja dijelova molekula, aktivira se
krajnji mehanizam, kontrolisana i programirana ćelijska smrt – apoptoza.
Cilj ovog mehanizma jeste da se ireverzibilno oštećene ćelije inkapsulišu
i odvoje od neoštećenih (Payne i sar. 1995). Svakako, apoptoza
može nastati i pod drugim uslovima, i smatra se jednim od glavnih mehanizama
kontrole rasta i proliferacije tkiva.
Otkrivanje i kvantifikacija oksidativnog stresa
Kao što je već navedeno, oksidativni stres predstavlja disbalans oksidanasa
i antioksidativne odbrane ćelije i cijelog organizma. On može nastati
kao posljedica povećanog unosa ili stvaranja oksidansa ili slabljenja
antioksidativne odbrane (zbog smanjenog unosa, nedovoljne sinteze ili
povećane potrošnje antioksidansa). S obzirom na visoku reaktivnost, u
dokazivanju i procjeni intenziteta oksidativnog stresa, direktno dokazivanje
oksidansa i antioksidansa rijetko se koristi u široj praksi (jer, pored
navedenog, zahtijeva sofisticiranu opremu i obučeno osoblje). Umjesto
toga, uglavnom se koriste indirektne metode, kao što je metoda koju navodi
Rosenkranz (1992), koja se zasniva na fluorescenciji boje dihlorofluorescina,
koja nastaje u reakciji njegovog nefluorescirajućeg diacetata i oksidansa).
Intenzitet fluorescencije može se mjeriti pomoću mikroskopa sa tamnim
poljem, čitača fluorescencije ili pomoću protočne citometrije, a smatra
se proporcionalnim intenzitetu oksidativnog stresa.
Mjerenje antioksidansa može biti fokusirano na određivanje ukupnog kapaciteta
antioksidativne mreže ili na određivanje aktivnosti i količine pojedinih
važnijih antioksidansa. Metode za određivanje ukupnog antioksidativnog
kapaciteta obuhvataju sljedeće metode: TAS (Total Antioxidant Status),
TEAC (Trolox Equivalent Antioxidant Capacity), TRAP (Total
Radical-trapping Antioxidant Parameter), FRAP (Ferric
Reducing-Antioxidant Power), kao i metodu ciklične voltametrije.
Većina ovih metoda dostupna je i komercijalno, u biohemijskim laboratorijama.
Sve ove metode imaju za cilj da odrede ukupni antioksidativni kapacitet
krvne plazme, izbjegavajući pri tome komplikovane metode određivanja aktivnosti
pojedinačnih antioksidanasa, a posebno njihovih međusobnih interakcija
(Cao i Prior, 1998; Niki and Noguchi, 2000). Od pomenutih metoda,
najčešće se koristi određivanje TAS, kao pokazatelj koji daje biološki
relevantnu informaciju o dinamičkoj ravnoteži između prooksidativnih i
antioksidativnih supstanci u krvnoj plazmi (Ghiselli, 2000).
Kada su u pitanju pojedinačni antioksidansi, najčešće se određuje količina
GSH i vitamina C. Mada postoji i niz drugih anitoksidativnih supstanci,
pomenuta dva su važna zbog svoje sposobnosti da regenerišu druge antioksidativne
supstance u prvobitno stanje. Prema Lykkesfeldtu (2002), kao direktni
biomarkeri oksidativnog stresa često se određuju i koncentracija vitamina
E i koenzima Q, kao lipofilnih antioksidanasa.
Visoko specifične metode za određivanje koncentracija GSH i vitamina C
se baziraju na upotrebi HPLC (tečne hromatografije visokih performansi),
kojom se određuje broj redoks-parova glutation-glutation disulfid i askorbat-dehidroaskorbat.
Jednostavnije, ali sa manjom specifičnošću redoks-status se može odrediti
i spektrofotometrijski. U oba slučaja, smanjena količina antioksidansa
i snižen odnos glutation-glutation disulfid i askorbat-dehidroaskorbat
redoks-parova tumači se kao indikator intenziviranja oksidativnog stresa.
Pored određivanja samih učesnika u redoks-reakcijama (bilo direktno ili
indirektno), procjena intenziteta oksidativnog stresa može se izvršiti
i na osnovu procjene intenziteta oštećenja glavnih ciljnih molekula oksidacije.
Oksidativna oštećenja DNK se, prema Collinsu (2004) najčešće otkrivaju
COMET metodom (gel elektroforeza koja semikvantitativno određuje broj
prekida lanca DNK u pojedinačnim ćelijama) ili se pomoću složene HPLC
ili masene spektrofotometrije određuje količina oksidovanih nukleotida
ili nukleozida. Markeri oštećenja DNK mogu se određivati u samom tkivu
(da se ustanovi stepen oštećenja na nivou ćelije) ili u mokraći (da bi
se ustanovila količina oksidovanih nukleotida i nukleozida, pri čemu postoji
sukob mišljenja da li povećanje njihove količine ukazuje na veći stepen
oksidacije ili veći intenzitet reparacije DNK putem isijecanja oštećenih
nukleotida). Proizvodi oksidacije proteina, karbonili nastali oksidacijom
aminokiselina, često se koriste kao markeri za procjenu oksidativnog stresa.
Njihova količina se određuje kolorimetrijskim metodama koje često pokazuju
ograničenu specifičnost. Uglavnom se koristi metoda sa dinitrofenilhidrazinom,
koji sa karbonilima gradi derivate koji imaju veliku moć apsorpcije svjetlosti
(Chevion i sar. 2000). Povećana apsorpcija svjetlosti (talasna
dužina preko 370 nm) se tumači kao indikator povišene oksidacije proteina.
U novije vrijeme dostupne su i druge metode, kojima se određuju specifični
produkti oksidacije pojedinih aminokiselina, ali se one rjeđe koriste
zbog specifične opreme i manjka senzitivnosti (jer se određuje samo nekoliko
od niza mogućih proizvoda reakcije).
Intenzitet oksidacije lipida može se utvrditi i procijeniti na više načina,
od kojih su u upotrebi najčešće određivanje koncentracija malondialdehida
(MDA) i izoprostana. MDA je međuproizvod razlaganja lipida putem oksidacije
i često se smatra indikatorom peroksidacije lipida u organizmu. Prema
Esterbaueru i Zollneru (1989), za određivanje koncentracije MDA se koristi
TBARS (Thiobarbiturate Acid Reactive Substances) metoda, kod
koje se MDA prvo vezuje sa tiobarbiturnom kiselinom, a zatim se količina
nastalog proizvoda određuje spektrofotometrijski.
Iako je upotreba MDA kao indikatora lipidne peroksidacije dosta kritikovana
(zbog niske specifičnosti i stvaranja dodatnih količina MDA tokom uzorkovanja
i reakcije sa TBA, kao i činjenice da MDA in vivo nastaje u još
nekoliko reakcija) ovo je jedan od najčešće korištenih markera oksidacije
lipida, uglavnom zbog svoje jednostavnosti određivanja. Novije verzije
TBARS metode mjere samo količinu čistog MDA-TBA2 derivata, putem HPLC
ili MDA direktno, bez vezivanja sa TBA (Lykkesfeldt, 2001). Prema
Morrowu (2000), određivanje MDA je jedna od boljih metoda za otkrivanje
oksidativnog oštećenja ćelije i pokazuje visoku korelaciju sa drugim (specifičnijim)
markerima, kao što su izoprostani. Prema novijim podacima (Lykkesfeldt,
2007), za određivanje izoprostana se danas, umjesto dosadašnje komplikovane
masene spektrofotometrije, koriste komercijalno dostupni ELISA kitovi.
Mliječne krave i oksidativni stres
Organizam visokomliječnih krava tokom proizvodno-reproduktivnog ciklusa trpi veliko metaboličko operećenje. Postoji čitav niz faktora koji mogu dovesti do nastanka oksidativnog stresa, među kojima je pomenuto metaboličko opterećenje jedno od najvažnijih. Od ostalih faktora kao važniji se navode ishrana (posebno ukoliko sadrži lako oksidišuće supstance, kao što su masne kiseline, koje se dodaju kako bi se popravila energetska vrijednost obroka), toplotni stres (koji intenzivira metaboličke procese, i time povećava produkciju reaktivnih metabolita različitih vrsta), i svakako metabolički status jedinke. U nastavku će biti dat pregled istraživanja različitih autora u vezi sa ovom tematikom, prema pomenutim faktorima.
Toplotni stres, nastao kao neminovna posljedica globalnog zagrijavanja, u novije vrijeme postaje sve veći problem za zdravlje ljudi i životinja, između ostalog i preko razvoja oksidativnog stresa i oštećenja ćelija. O povezanosti ovih pojava sproveden je niz istraživanja, a posebno interesantne su se pokazale mliječne krave. Bellò-Klein i sar. (2000) su u svome istraživanju utvrdili da su temperatura i relativna vlažnost vazduha faktori koji imaju presudan uticaj na razvoj oksidativnog oštećenja organizma tokom nastanka toplotnog stresa, što je, generalno, vezano za sezonu, odnosno godišnje doba. Ovi autori ne isključuju mogućnost da, pored pomenutih, postoje još neki faktori koji mogu doprinijeti razvoju oksidativnog stresa.
Bernabucci i sar. (2002) su ukazali na povezanost između ambijentalne temperature i pojave oksidativnog stresa mliječnih krava, odnosno promjene koncentracije indikatora oksidativnog stresa (ROM u krvnoj plazmi, TBARS, glutation-peroksidaza i tiolne grupe u krvnoj plazmi i eritrocitima, superoksid-dismutaza u eritrocitima). Oni su ispitivali oksidativni status krava koje su u peripartalnom periodu držane u temperaturno povoljnim i uslovima toplotnog stresa (proljetni i ljetni period) i ustanovili da je u uslovima toplotnog stresa koncentracija indikatora oksidativnog stresa bila viša u odnosu na drugu grupu krava. Prateći markere oksidativnog stresa u eritrocitima i krvnoj plazmi, ovi autori nisu ustanovili statistički značajnu razliku između grupa krava, kada su u pitanju bili markeri u plazmi, dok je kod eritrocitnih markera ta razlika postojala. Zaključak njihovog istraživanja bio je da temperaturni uslovi koji mogu dovesti do nastanka toplotnog stresa mogu, između ostalog, imati za posljedicu i nastanak oksidativnog oštećenja organizma. Pri tome su eritrocitni markeri označeni kao pouzdaniji indikatori oksidativnog oštećenja organizma u odnosu na markere oksidativnog stresa koji se uobičajeno određuju u krvnoj plazmi. Njihova pretpostavka je da je istovremena povećana aktivnost eritrocitnih indikatora oksidativnog stresa indirektni kompenzujući odgovor ćelija na povećanu oksidativnu aktivnost tokom razvoja toplotnog stresa, odnosno povećani utrošak antioksidativnih supstanci. Kao potencijalni razlog za izostanak statistički značajne razlike u vrijednostima markera oksidativnog oštećenja u krvnoj plazmi u vezi sa sezonom, ovi autori navode kombinaciju umjerenog toplotnog stresa i pojačane antioksidativne aktivnosti krvne plazme, pozivajući se na istraživanje Frei-a i sar. (1998). Kao razlog za veću pouzdanost eritrocitnih markera oksidativnog stresa i veću osjetljivost eritrocita na oksidativno oštećenje Clemens i Waller (1987) navode visok sadržaj polinezasićenih masnih kiselina u ćelijskoj membrani eritrocita, koja je permanentno izložena visokim koncentracijama kiseonika i reaktivnom gvožđu iz hemoglobina. Zbog toga Kusmic i sar. (2000) i Alicigüzel i sar. (2001) označavaju eritrocite kao adekvatan model za ispitivanje oksidativnog stresa. Još jedan od razloga veće osjetljivosti eritrocita na oksidativno oštećenje tokom toplotnog stresa jeste povećan parcijalni pritisak kiseonika u krvi, nastao zbog ubrzanog i površnog disanja jedinke, pri čemu se unosi veća količina kiseonika, a izbacuje manja količina ugljen-dioksida u odnosu na uobičajene fiziološke vrijednosti. Di Guiseppi i sar. (1984) navode da u takvim uslovima povećan sadržaj visoko nezasićenih masnih kiselina u membrani eritrocita i prisustvo visoko reaktivnog jona gvožđa u sastavu hemoglobina mogu imati za posljedicu povećano stvaranje slobodnih radikala i slabljenje antioksidativne odbrane eritrocita.
Bernabucci i sar. (2002) kao indikator adaptacione sposobnosti krava na uslove toplotnog stresa označavaju povišenje aktivnosti superoksid-dismutaze i koncentracije tiolnih grupa u eritrocitima, pozivajući se na navode Harmona i sar. (1997) i Calamarija i sar. (1999) koji su ustanovili smanjenu antioksidativnu aktivnost u krvnoj plazmi krava tokom sredine laktacije, kada su one bile izložene uslovima toplotnog stresa. Isti autori navode rezultate Trouta i sar. (1998) koji su ustanovili da kratkotrajno izlaganje visokim ambijentalnim temperaturama nema uticaj na koncentracije liposolubilnih antioksidanasa (α-tokoferola, β-karotina, retinola i retinil-palmitata) u krvi i koncentracije MDA u mišićima, iz čega se može izvesti zaključak da je za izazivanje oksidativnog stresa potrebno da organizam bude duže vrijeme izložen nepovoljnim ambijentalnim uslovima.
Noble i sar. (1976) su ustanovili da se udio linolenske kiseline u frakcijama holesteril estara smanjuje ukoliko su goveda izložena uslovima toplotnog stresa. Ovi autori se u svome radu pozivaju na istraživanje Kiupera i sar. (1971) koji su ustanovili smanjenje koncentracije linolenske kiseline u membranama eritrocita hrčkova izloženih toplotnom stresu. Jedan od zaključaka rada Bernabuccija i sar. (2002) jeste da je poremećen antioksidativni status eritrocita kod krava izloženih toplotnom stresu, manifestovan preko povećanja TBARS, između ostalog posljedica smanjenog sadržaja linolenske kiseline u njihovoj membrani, koji ima za posljedicu njenu veću fragilnost.
Hormonalni status jedinke, pogotovo kada su u pitanju metabolički aktivni hormoni, takođe može biti faktor koji utiče na nastanak oksidativnog stresa. Alicigüzel i sar. (2001) su postavili hipotezu da hipermetabolički efekti tireoidnih hormona u uslovima hipertireoidizma mogu biti uzrok oksidativnog oštećenja eritrocita. Kada su u pitanju visokomliječne krave, ovaj faktor se može zanemariti, jer je poznato da krave u uslovima toplotnog stresa imaju niži nivo tireoidnih hormona, i uopšte snižen nivo metabolizma, što potvrđuju i podaci koje navode Webster (1991) i Nardone i sar. (1997). Kada je u pitanju peripartalni period i rana laktacija, situacija je slična, jer i tada fiziološki dolazi do snižavanja bazalnog nivoa tireoidnih hormona. Dodatni razlog je to što se kod krava izrazito visoke mliječnosti relativno često pojavljuje subklinički hipotireoidizam, kao odraz neadekvatne adaptacije organizma na metaboličko opterećenje izazvano visokom mliječnošću (Šamanc i sar., 2010).
Niz autora (Grummer, 1993; Goff and Horst, 1997; Drackley, 1999) navodi da je peripartalni (tranzicioni) period izrazito važan momenat za zdravlje i proizvodne sposobnosti mliječnih krava, i da su one u tom periodu izložene ogromnim fiziološkim izazovima i metaboličkom stresu. Metabolički procesi u peripartalnom periodu nalaze se pod kontrolom veoma suptilnih homeoretskih kontrolnih mehanizama, kako bi se rad tkiva i organa prilagodio potrebama organizma u datom periodu (Vernon, 2002). Intenzivna lipomobilizacija i ogromne promjene u prometu energije do kojih dolazi u relativno kratkom periodu oko teljenja predstavljaju značajan izvor oksidanasa i potencijalni okidač za nastanak oksidativnog stresa. Istraživanjima je utvrđeno da postoji čitav niz faktora koji utiču na pojavu poremećaja zdravlja krava u peripartalnom periodu, među kojima su svakako najpoznatiji tjelesna kondicija, ishrana i metabolički status. Veza između tjelesne kondicije, odnosno stepena ugojenosti, i pojave poremećaja zdravlja opisana je od strane velikog broja autora i ustanovljena je jasna pozitivna korelacija između ove dvije pojave. Hipotezu o povezanosti oksidativnog stresa i poremećaja zdravlja krava u peripartalnom periodu navodi više autora koji su se bavili razmatranjem etiopatogeneze najčešćih poremećaja metabolizma u ovom periodu (Harrison i sar. 1984; Smith i sar. 1984; Gröhn i sar. 1989; Lomba i sar. 1996). Miler i sar. (1993) su pratili uticaj reaktivnih metabolita kiseonika na pojavu najučestalijih bolesti krava, između ostalog i mastitisa, i ustanovili jasnu pozitivnu korelaciju. Prema ovim autorima, poremećaj homeostaze, nastao zbog intenzivnog metaboličkog opterećenja organizma visokom mliječnošću, ima za posljedicu pojavu oksidativnog stresa, koji ima i direktni i indirektni uticaj na zdravlje životinja. Bernabucci i sar. (2002) toplotni stres označavaju kao faktor koji potencira već postojeće oksidativno oštećenje organizma kod krava u peripartalnom periodu.
Veza oksidativnog stresa i tjelesne kondicije u peripartalnom periodu detaljnije je obrađena u radu Bernabuccija i sar. (2005), koji su u svom istraživanju ustanovili da postoji korelacija između oksidativnog i energetskog statusa krava, u smislu da su krave koje su prije teljenja bile ugojene i tokom postpartalnog perioda više gubile na tjelesnoj masi bile znatno osjetljivije na oksidativni stres od životinja optimalne tjelesne kondicije. Između gubitka tjelesne mase i intenziteta oksidativnog oštećenja ustanovljena je pozitivna korelacija. Kao parametri za procjenu oksidativnog statusa krava i intenziteta oksidativnog stresa u ovom istraživanju uzeti su sadržaj ROM, TBARS, SH- (tiolnih) grupa i aktivnost glutation-peroksidaze u krvnoj plazmi, aktivnost glutation-peroksidaze i superoksid-dismutaze i sadržaj tiolnih grupa u eritrocitima. Rezultati koje su dobili na uzorcima uzetim u postpartalnom periodu pokazali su smanjenje aktivnosti superoksid-dismutaze i koncentracije tiolnih grupa u krvnoj plazmi i eritrocitima, kao i povišenje koncentracije ROM, TBARS i aktivnosti glutation-peroksidaze u krvnoj plazmi u odnosu na vrijednosti dobijene prepartalno. Nalaz je ukazivao na postojanje oksidativnog stresa zbog pojačanog stvaranja oksidansa tokom intenzivne lipomobilizacije i slabljenja kapaciteta antioksidativne odbrane. Metabolički profil ispitanih krava ukazivao je na povećan intenzitet lipomobilizacije kod krava koje su prepartalno bile ugojene (povišen nivo BHBA i NEFA), pri čemu je postojala statistički značajna razlika u gubitku tjelesne mase ugojenih krava od kasnog graviditeta do 30 dana poslije teljenja u odnosu na krave optimalne tjelesne kondicije i krave sa OTK ispod optimalne.
Bernabucci i sar. (2005) su ustanovili da je antioksidativni status krava u peripartalnom periodu oslabljen, zbog povećane potrošnje antioksidansa. U ovoj fazi proizvodno-reproduktivnog ciklusa može doći do pojave oksidativnog stresa, što se može zaključiti na osnovu povećanja koncentracije TBARS i glutation-peroksidaze u krvnoj plazmi. Iz pomenutog nalaza ovi autori postavljaju hipotezu da metaboličko i endokrino prilagođavanje, koje organizam sprovodi da bi se prilagodio potrebama fetusa i sekrecije mlijeka u peripartalnom periodu, može imati za posljedicu oksidativni stres.
Miller i sar. (1993) navode da prije teljenja dolazi do smanjenja nivoa ROM, jer je antioksidativna odbrana u ovoj fiziološkoj fazi snažnija. Hermes-Lima i sar. (1998) iznose mišljenje da je aktivacija antioksidativnih mehanizama, koja dovodi do smanjenja produkcije i oslobađanja slobodnih radikala, ustvari predstavlja pripremni mehanizam za odbranu od oksidativnog stresa, uzrokovan fiziološki stresnim situacijama, kao što su porođaj i laktacija. Miller i sar. (1994) smatraju da je povećana metabolička aktivnost nakon teljenja jedan od razloga za pojačan rad transportnog lanca elektrona i produkciju ROM. Barja de Quiroga i sar. (1992) smatraju da je produkcija endogenih antioksidanasa pod kontrolom homeostatskih mehanizama, te da se povećava paralelno sa izlaganjem dejstvu oksidanasa.
Kowaliszyn i sar. (2008) su istraživali postojanje korelacije između koncentracije MDA u krvnoj plazmi i fiziološkog stanja krava, i korelaciju između koncentracije MDA u plazmi i broja somatskih ćelija u mlijeku. Njihovo istraživanje obuhvatilo je podjednak broj krava pred zasušenje i krava mjesec dana nakon teljenja. Ovi autori nisu ustanovili statistički značajnu razliku između posmatranih grupa krava u odnosu na njihov fiziološki status, dok je unutar grupe krava pred zasušenje ustanovljena statistički značajna razlika između grla sa različitim brojem somatskih ćelija, pri čemu je viša koncentracija MDA ustanovljena kod grla sa manjim brojem somatskih ćelija u mlijeku. Izostanak statistički značajnih razlika u koncentraciji MDA u krvnoj plazmi između krava različitog fiziološkog statusa može se protumačiti izborom stadijuma laktacije u kome su uzorci uzimani. Naime, kod grupe krava koje su bile u laktaciji već mjesec dana je prošao najkritičniji period od nekoliko dana oko samog teljenja, kada je produkcija slobodnih radikala i oštećenje ćelija, a sa njima i koncentracija MDA u plazmi na najvišem nivou.
Castillo i sar. (2005) su ispitivali povezanost fiziološkog statusa krava i pojave oksidativnog stresa, kao i povezanost markera oksidativnog stresa i vrijednosti parametara metaboličkog profila, kao uobičajenog dijagnostičkog testa za otkrivanje poremećaja zdravlja životinja. Ovi autori su se bazirali na procjenu oksidativnog oštećenja organizma preko praćenja koncentracije MDA u krvnoj plazmi, kao pokazatelja peroksidacije lipida i TAS. Ogledne životinje bile su u periodu kasne laktacije i u peripartalnom periodu (praćene su od deset nedelja prije do dvije nedelje nakon teljenja, a uzorci krvi su uzimani jednom nedeljno). Obe grupe krava hranjene su kompletnim obrokom, prilagođenim potrebama životinja u datoj fiziološkoj fazi, i nisu pokazivale poremećaje zdravlja, što je važno naglasiti, s obzirom da se većina radova na ovu temu bazira na životinjama sa klinički manifestnim poremećajima zdravlja. Posebnu važnost u ovom istraživanju ima sastav obroka, koji je bio koncipiran tako da u potpunosti prati nutritivne i energetske potrebe životinja, te su se svi parametri metaboličkog profila tokom cijelog istraživanja održali unutar fizioloških granica. Parametri metaboličkog profila se, u principu, nisu značajnije razlikovali između različitih grupa krava, a statistički značajne razlike između grupa ustanovljene su kod koncentracija triglicerida, holesterola, uree i AST. Unutar grupe krava u peripartalnom periodu ustanovljene su statistički značajne razlike u koncentracijama triglicerida, holesterola, ukupnih proteina, uree, kreatinina i AST. Ovakve razlike u vrijednostima parametara metaboličkog profila su, na osnovu empirijskih podataka, i očekivane. Oksidativni status ispitanih krava pokazivao je statistički značajne razlike kada je u pitanju TAS (i između grupa, i unutar grupe krava u peripartalnom periodu), dok koncentracija MDA nije pokazivala statistički značajna odstupanja. Koncentracija MDA nije pokazivala jasan trend tokom peripartalnog perioda, dok je TAS imao jasan trend povećanja vrijednosti sve do nedelju dana pred teljenje, da bi nakon toga opadao, ali je do kraja istraživanja ostao na vrijednostima iznad prepartalnih. Pik vrijednosti MDA registrovan je nedelju dana nakon teljenja, ukazujući na intenzivne metaboličke procese u kratkom vremenskom periodu oko teljenja. Ovaj nalaz je u skladu sa rezultatima do kojih su došli Turk i sar.(2004), koji su, u cilju procjene oksidativnog statusa mliječnih krava u ranom postpartalnom periodu, pratili aktivnost serumske paraoksonaze.
Da bi utvrdili uticaj stadijuma laktacije, odnosno graviditeta na vrijednosti parametara metaboličkog profila i oksidativnog stresa, autori su krave grupisali u četiri grupe (kasna laktacija, rano zasušenje, kasno zasušenje, rana laktacija) i ustanovili statistički značajne razlike u koncentracijama holesterola između svih grupa, uree između grupa krava u zasušenju međusobno, krava u kasnom zasušenju i ranoj i kasnoj laktaciji, kreatinina između svih grupa krava, alkalne fosfataze između grupa krava unutar perioda zasušenja i u pogledu ukupnog antioksidativnog statusa između krava u ranom zasušenju i ranoj laktaciji. Kada je u pitanju poređenje sa podacima drugih autora, Castillo i sar. ne navode radove sa sličnom metodologijom, te je tumačenje rezultata izvršeno u odnosu na literaturne podatke o uticaju antioksidativnog statusa na homeoretske mehanizme koji se aktiviraju u peripartalnom periodu i o korelaciji između metaboličkih i pokazatelja oksidativnog statusa jedinke. Kada je u pitanju ukupni antioksidativni status, autori navode da dobijeni rezultati kod krava u kasnoj laktaciji i graviditetu nisu u skladu sa njihovim prethodnim istraživanjima, sprovedenim na kravama na vrhuncu laktacije (Castillo i sar., 2003). Dobijeni podaci su, prema ovim autorima, u skladu sa rezultatima koje su dobili Watcher i sar. (1999), koji su ustanovili progresivno slabljenje antioksidativne aktivnosti krvne plazme sa napredovanjem laktacije, koje je najvjerovatnije u vezi sa izlučivanjem liposolubilnih antioksidansa preko mlijeka.
Ispitivanje korelacije između vrijednosti metaboličkih i parametara antioksidativnog statusa krava u različitim stadijumima laktacije, odnosno graviditeta, pokazalo je da je kod krava u kasnoj laktaciji pozitivna korelacija postojala između TAS i koncentracije albumina, a kod krava u ranoj laktaciji između koncentracije MDA i triglicerida. U grupi krava u kasnom zasušenju ustanovili su pozitivnu korelaciju između koncentracija MDA i NEFA i TAS i koncentracije albumina. Kod krava u ranom zasušenju ovi autori ustanovili su negativnu korelaciju između koncentracija glukoze i MDA i koncentracije MDA i aktivnosti alkalne fosfataze, a kod krava u kasnom zasušenju između TAS i koncentracije holesterola u krvi.
Castillo i sar. (2005) su, prateći povezanost metaboličke aktivnosti i oksidativnog statusa krava ustanovili jasnu korelaciju, u odnosu na fiziološko stanje krava. Kod krava koje u periodu kasne laktacije nisu ispoljavale statistički značajne razlike u vrijednostima parametara metaboličkog profila u odnosu na gravidne krave, koncentracija MDA nije bila u korelaciji sa bilo kojim parametrom metaboličkog profila. Izostanak korelacije bio je u skladu sa vrijednostima praćenih parametara koje govore u prilog "normalnoj" metaboličkoj aktivnosti, odnosno nepostojanju oksidativnog stresa. Kod krava u peripartalnom periodu ustanovljeno je postojanje korelacije između pojedinih parametara, što ukazuje na povezanost metaboličke aktivnosti i produkcije lipoperoksida. Tako je kod krava u kasnom zasušenju ustanovljena negativna korelacija između lipoperoksida sa jedne i glikemije i aktivnosti alkalne fosfataze sa druge strane, dok je dvije nedelje prije i neposredno nakon teljenja koncentracija lipoperoksida pokazivala jaku pozitivnu korelaciju sa indikatorima metabolizma lipida. S obzirom na dokazanu pozitivnu korelaciju između NEFA i triglicerida u krvi (Bell, 1995; Vanderhaar i sar., 1999) i pojačan oksidativni metabolizam u peripartalnom periodu, logično je očekivati pojačanu peroksidaciju oslobođenih masnih kiselina i nastanak lipoperoksida (Kankofer, 2001).
Jedan od zaključaka ovog rada, na koji ukazuju velike varijacije u vrijednostima praćenih parametara unutar grupa, jeste da, pored uticaja fiziološke faze (odnosno metaboličkog opterećenja), na oksidativni status jedinke u velikoj mjeri utiču i individualne varijacije u aktivnosti homeostatskih i homeoretskih mehanizama, kao i niz drugih faktora koje tek treba istražiti. Jedan od tih faktora je svakako napor pri teljenju, odnosno pojačana mišićna aktivnost (autori se pozivaju na istraživanja Chiaradie i sar. iz 1998, koji su, ispitujući efekte kratkotrajnog intenzivnog opterećenja, kod trkačkih konja ustanovili značajno povećanje potrošnje kiseonika u mišićima tokom trke, koje je izvor povećane oksidativne aktivnosti). Poznata je činjenica da je svaki porođaj specifičan u pogledu napora, odnosno mišićnog rada koji organizam majke ulaže da bi se uspješno dovršio. Kao potencijalne faktore bi trebalo uzeti u obzir dužinu trajanja porođaja i eventualnu pomoć pri teljenju, na koje u ovom istraživanju nije obraćena pažnja.
Ishrana, odnosno sastav obroka, takođe u velikoj mjeri može da utiče na oksidativni status, kao što je ranije rečeno. U istraživanju koje su sproveli Castillo i sar. (2005) najniže registrovane vrijednosti TAS koincidirale su sa početkom zasušenja (oko deset nedelja pred očekivano teljenje) i smanjenjem učešća vitaminsko-mineralnih dodataka u obroku, kao izvora egzogenih antioksidansa. Ipak, u periodu do teljenja došlo je do postepenog oporavka i jačanja antioksidativne aktivnosti plazme, što se može pripisati pojačanoj sintezi endogenih antioksidansa, prije svega albumina. Da se ovaj oporavak zasniva prvenstveno na produkciji endogenih antioksidansa govori i podatak koji navode Goff i Horst (1997), da se pred samo teljenje koncentracija pojedinih antioksidansa (posebno vitamina E) smanjuje do granice deficita. Curtis (1997) navodi tvrdnju da se stresom uslovljena pojačana antioksidativna aktivnost krvne plazme, kao fiziološki odbrambeni mehanizam, može očekivati u periodima kada je opasnost od oksidativnog oštećenja organizma izražena, i da je taj efekat nije u vezi sa snabdjevenošću vitaminima i mineralima, kao poznatim egzogenim antioksidansima (Brzezinska-Slebodzinska i sar., 1994). Rezultati koje su dobili Castillo i sar. (2005) pokazali su da je TAS dostigao pik neposredno nakon teljenja, istovremeno sa pikom MDA, što govori da je antioksidativni sistem organizma, potpomognut vitaminima i mineralima dodatim u obrok, u stanju da se efikasno izbori sa nastalim lipoperoksidima tokom kritičnog ranog postpartalnog perioda, odnosno potencijalnim izvorom oksidativnog stresa (Miller, 1993; Castillo i sar., 2001). Castillo i sar. (2001) su u svome radu potvrdili jasnu povezanost između TAS i albumina, kao komponente sa velikim antioksidativnim kapacitetom, ukazujući pri tome na posebnu važnost albumina u periodu kasnog zasušenja, kada životinje ne dobijaju vitaminsko-mineralne dodatke, a dolazi do intenziviranja oksidativnih procesa i stvaranja većih količina MDA. Važnost albumina kao komponente antioksidativne odbrane dobija na značaju ukoliko se uzme u obzir negativna korelacija između TAS i holesterolemije, jer albumini utiču na smanjenje potrošnje citohroma P-450, neophodnog za reakciju razlaganja holesterola, u kojoj nastaju reaktivni metaboliti kiseonika, te tako potpomaže održavanje ravnoteže između prooksidativnih i antioksidativnih supstanci.
Generalni zaključak rada Castillo i sar. (2005) jeste da, pored fiziološkog statusa, postoji još čitav niz nedovoljno razjašnjenih faktora koji mogu da utiču na oksidativnu aktivnost u peripartalnom periodu, te da je na njihovom otkrivanju i razjašnjavanju njihovih međusobnih interakcija neophodno dalje raditi. Kao uopštenu preporuku i mjeru prevencije nastanka oksidativnog stresa u peripartalnom periodu, ovi autori navode obezbjeđenje dovoljne količine minerala i vitamina u obroku za krave u periodu zasušenja, čak i u slučaju da ne pokazuju simptome oksidativnog oštećenja organizma.
Da bi utvrdili oksidativni status krava u različitim fazama laktacije, Castillo i sar. (2006) su sproveli istraživanje na kravama u kasnoj laktaciji (oko sedmog mjeseca laktacije) i kravama u početnoj fazi laktacije (ove životinje praćene su od prve do osme nedelje nakon teljenja). Sve životinje hranjene su kompletnim TMR obrokom, prilagođenim njihovim energetskim i nutritivnim potrebama. Slično prethodnom istraživanju (Castillo i sar., 2005), i ovdje su paralelno praćeni parametri oksidativnog statusa (koncentracija MDA i TAS) i uobičajeni parametri metaboličkog profila (glikemija, koncentracije uree, triglicerida, albumina, NEFA, ukupnih proteina, kreatinina, aktivnost AST i alkalne fosfataze). Statistički značajne razlike nisu ustanovljene između grupa krava i unutar grupe krava u ranoj laktaciji kada su u pitanju glikemija, koncentracija albumina i uree i aktivnost AP. Koncentracija NEFA pokazivala je statistički značajnu razliku unutar grupe krava u ranoj laktaciji (druga i osma nedelja laktacije međusobno), a takođe i između obe grupe krava (druga, četvrta i osma nedelja u odnosu na krave u kasnoj fazi laktacije). Ukupni proteini su se statistički značajno razlikovali kod krava u ranoj laktaciji (prva i druga nedelja laktacije) i između grupa krava (druga nedelja laktacije u odnosu na krave u kasnoj laktaciji). Koncentracija kreatinina bila je statistički značajno viša kod krava u ranoj laktaciji (prva i druga nedelja) u odnosu na krave u kasnoj laktaciji, da bi se, sa vrhuncem laktacije, izjednačila kod obe grupe krava. Aktivnost AST nije pokazivala statistički značajna odstupanja tokom laktacije, sem u četvrtoj nedelji nakon teljenja, dok je prosječna aktivnost bila veća tokom rane u odnosu na kasnu laktaciju. Prosječna koncentracija MDA nije pokazivala statistički značajna odstupanja između grupa krava, i nije bilo jasnog trenda njenog kretanja tokom rane laktacije. Ipak, posebno interesantan nalaz bila je činjenica da je najveća prosječna vrijednost koncentracija MDA, a istovremeno i najviša standardna greška (koja je bila uzrok izostanka statističke značajnosti) registrovana upravo u vrijeme kada su, prema Grummeru (1993), krave najosjetljivije na nastanak metaboličkih poremećaja. U skladu sa ovim nalazom, autori ukazuju da je potrebno razmotriti uzroke individualnih varijacija u oksidativnom statusu pojedinih krava, te navode da su najviše vrijednosti koncentracije MDA koincidirale sa najvećim varijacijama u dnevnom unosu hrane. U prilog navode istraživanje Ingvartsena i Andersena (2000) koji su ustanovili značajne individualne varijacije u dnevnom unosu hrane, koje su u vezi sa nizom još uvijek nerazjašnjenih faktora, kao što su sastav obroka i promjene u homeostazi. Takođe, autori pokušavaju da smanjen unos hrane ne tumače kao neadekvatan adaptivni odgovor organizma, već kao organizovanu strategiju kojom organizam pokušava da podrži preživljavanje u energetski nepovoljnim uslovima. Ovo objašnjenje je, po njima, u skladu sa teorijom koju su postavili Ketelaars i Tolkamp (1996), koji tvrde da životinje unose neto energiju hrane u količini koja obezbjeđuje najoptimalniju potrošnju kiseonika i minimizira stvaranje slobodnih radikala.
TAS takođe nije pokazivao statistički značajne razlike između grupa, ali je unutar grupe krava u ranoj laktaciji pokazivao jasan trend opadanja sa odmicanjem laktacije. Naviša vrijednost TAS registrovana je nedelju dana nakon teljenja, zajedno sa najvišom vrijednošću koncentracije MDA. Kombinacija kretanja vrijednosti TAS i koncentracije MDA ukazuje da se oba parametra zajedno povećavaju i smanjuju, ukazujući da je na vrhuncu laktacije produkcija lipoperoksida na niskom nivou, najvjerovatnije kao posljedica adaptacionih procesa, započetih tokom ranog postpartalnog perioda. Vrijednosti TAS ustanovljene u ovom istraživanju u svim fazama laktacije bile su niže u odnosu na prethodna istraživanja istih autora, sprovedena na kravama na vrhuncu laktacije (Castillo i sar., 2003). Ustanovljeni pad antioksidativne aktivnosti sa odmicanjem laktacije u ovom istraživanju bio je u skladu sa nalazima Watchera i sar. (1999), koji su ustanovili da se antioksidativna aktivnost plazme smanjivala sa laktacijom, tumačeći to izlučivanjem liposloubilnih antioksidansa putem mlijeka.
Veoma interesantan nalaz koji se navodi u oba rada Castillo i sar. (2005, 2006) jeste uočena stabilnost glikemije, koja nije bila u skladu sa očekivanim varijacijama tokom različitih fizioloških faza, a koja se može dovesti u vezu sa načinom ishrane posmatranih krava. Naime, prepartalni obrok ovih krava sadržavao je više proteina u odnosu na preporuke NRC (1989), što je imalo za posljedicu podizanje glikemije (odnosno njeno održavanje unutar fizioloških granica). Ovaj nalaz potvrđuju i rezultati do kojih su došli Putnam i Varga (1998), koji su zaključili da do podizanja glikemije dolazi zbog većeg priliva glukoneogenih aminokiselina. Pored povećane količine proteina, obrok krava koje su pratili Castillo i sar. (2005, 2006) sadržavao je i veću količinu NFC (non-fiber carbohydrates, ne-vlaknasti ugljeni hidrati, odnosno skrob i drugi lako svarljivi ugljeni hidrati) u odnosu na preporuke NRC, tokom čije fermentacije je nastajala veća količina propionata, glavnog prekursora glukoze krvi (Minor i sar., 1998). Zaključak ovih radova jeste da je kombinacija povećane količine proteina i NFC rezultovala očuvanjem glikemije, iako je na samom početku laktacije registrovan njen pad (što je prema Bellu, 1995, u vezi sa sintezom laktoze), ali je vrijednost ipak ostala unutar fizioloških granica. U ranom postpartalnom periodu (prve dvije nedelje nakon teljenja) je registrovan uobičajeni lipidni status, opisan od strane Marcosa i sar. (1990), a koji jasno ukazuje na lipomobilizaciju, kao posljedicu specifične hormonalne konstelacije u ovom periodu (pojačana sekrecija hormona rasta i hormonski indukovana rezistencija masnog tkiva na insulin), koja potencira lipolizu i mobilizaciju NEFA, kako bi se zadovoljile energetske potrebe mliječne žlijezde (Grum i sar., 1996; Minor i sar., 1998; Doepel i sar., 2002). Pojačano snabdijevanje krava energijom i proteinima u prepartalnom periodu, prema navodima Parka i sar. (2002) može biti ključni momenat u adaptaciji metabolizma na potrebe u peripartalnom periodu. Smanjen unos hrane, koji je uobičajen u ranom postpartalnom periodu (Agenäs i sar., 2003), registrovan je i u ovom istraživanju, pri čemu nije postojala statistički značajna razlika u količini unijete hrane. Povećanje koncentracije ukupnih proteina, koje je registrovano u ranom postpartalnom periodu ukazuje na mobilizaciju tkivnih proteina, kao i na pojačanu snabdjevenost proteinima u obroku. Kako navode Vanderhaar i sar. (1999), ovi proteini služe kao izvor aminokiselina za metabolizam u mliječnoj žlijezdi i glukoneogenezu u jetri, procese čije je odvijanje tokom najranije faze laktacije kritično. Sa mobilizacijom tkivnih proteina, prvenstveno iz skeletnih mišića, dolazi i do povećanja koncentracije kreatinina. Pojačana sintetska aktivnost jetre je svakako u vezi sa mobilizacijom pomenutih tjelesnih rezervi (Park i sar., 2002), na šta ukazuje statistički značajno, ali ne i patološko povišenje aktivnosti AST.
Objašnjenje za pad koncentracije NEFA nakon druge nedelje laktacije najvjerovatnije je odraz značajnog izlučivanja NEFA putem mliječne masti (Bell, 1995), promjena endokrine konstelacije (prema nalazima Ingvartsena i Andersena, 2000, i Doepela i sar., 2002, tokom druge nedelje po teljenju dolazi do pada koncentracije hormona rasta i rasta koncentracije insulina) i povećanja dnevnog unosa hrane i energetskog bilansa (Vanderhaar i sar., 1999). Minor i sar. (1998) navode da do opadanja koncentracije NEFA može doći i na račun povećanog unosa NFC, kao prekursora propionata i glukoze krvi, sa odgovarajućim antilipolitičkim efektom. Prema nalazima Xua i sar. (1998), promjene u aktivnosti AST koje postoje u ovom periodu, posljedica su povećanog unosa proteina putem obroka i pojačanog katabolizma tkivnih proteina.
Na osnovu svog istraživanja i navoda iz literature, Castillo i sar. (2006) navode da je koncentracija MDA u krvnoj plazmi tokom laktacije ukazuje na sljedeće događaje: tokom rane laktacije u organizmu nastaje značajna količina slobodnih radikala, koji mogu dovesti do peroksidacije tkivnih lipida; pomenuti efekat vezan je za intenzitet metaboličkih promjena, koje su pod kontrolom endokrinih mehanizama, i dešava se na samom početku laktacije; pojačana oksidativna aktivnost postoji veoma kratak vremenski period. Ipak, ovi autori navode da nalaze povišene koncentracije MDA i TAS treba pažljivo tumačiti, zajedno sa rezultatima drugih ispitivanja (prije svega parametara metaboličkog profila), te da njihove vrijednosti mogu u značajnoj mjeri doprinijeti formiranju slike o homeostatskim i homeoretskim procesima koji se tokom tranzicionog perioda odvijaju u organizmu visokomliječnih krava. Postojanje značajnih individualnih razlika u vrijednostima pojedinih parametara oksidativnog statusa (prije svega MDA), kao i donekle nejasna korelacija sa pojedinim parametrima metaboličkog profila, prema ovim autorima, treba da budu osnova za dalja istraživanja, koja će u potpunosti razjasniti učešće ovih faktora u formiranju i održavanju oksidativnog statusa životinja.
Zaključak
Oksidativni stres je stanje koje nastaje kada antioksidativna odbrana
organizma nije više u mogućnosti da se izbori sa produkcijom slobodnih
radikala, prije svega reaktivnih metabolita kiseonika. Organizam ima više
nivoa antioksidativne odbrane, koji se aktiviraju tokom oksidativnog oštećenja
tkiva, ćelija i cijelog organizma, i onemogućavaju nastanak značajnijih
oštećenja. Nakon niza istraživanja, određeni su parametri koji ukazuju
na oksidativni status jedinke, te definisane metode za njihovo određivanje.
Visokomliječne krave, naročito tokom tranzicionog perioda, su veoma interesantne
kada je u pitanju oksidativni stres, te je u skladu sa time sproveden
niz istraživanja. Pored ogromnog metaboličkog opterećenja nastalog prilikom
prelaska iz perioda zasušenja u period rane laktacije, potencijalni izvor
oksidansa za ovu kategoriju životinja su još ishrana, toplotni stres i
niz drugih, manje ili više razjašnjenih faktora. U radu je dat pregled
dostupnih istraživanja na ovu temu, sa akcentom na korelaciju između parametara
metaboličkog profila i oksidativnog statusa jedinke.
Literatura
- Agenas, S., Burstedt, E., Holtenius, K. (2003): Effects of feeding intensity during the dry period. 1. Feed intake, body weight, and milk production, J. Dairy Sci. 86, 870-882
- Aliciguzel, Y., Ozdem. S.N., Ozdem, S.S., Karayalcin, U., Siedlak, S.L., Perry, G., Shith, M.A. (2001): Erithrocyte, plasmam and serum antioxidant activities in untreated toxic multinodular goiter patients, Free Radic. Biol. Med. 30:665-670
- Barja de Quiroga, G., Lopez-Torres, M., Perez-Campo, R. (1992): Relationship between antioxidants, lipid peroxidation and aging, EXS 62:109-123
- Bell, A.W. (1995): Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation, Journal of Animal Science 73:2804-2819
- Bello-Klein, A., Morgan-Martins, M.I., Barp, J., Llesuy, S., Bello, A.A., Singal, P.K. (2000): Circaanual changes in antioxidants and oxidative stress in the heart and liver in rats,Comp. Biochem. Physiol. C 126:203-208
- Bernabucci, U., Ronchi, B., Lacetera, N., Nardone, A. (2002): Markers of Oxidative Status in Plasma and Ertithrocytes of Transition Dairy Cows During Hot Season, J. Dairy Sci. 85:2173-2179
- Bernabucci, U., Ronchi, B., Lacetera, N., Nardone, A. (2005): Influence of Body Condition Score on Relationships Between Metabolic Status and Oxidative Stress in Periparturient Dairy Cows, J. Dairy Sci. 88:2017-2026
- Brzezinska-Slebodzinska, J.K., Miller, J.K., Quigley, J.D., Moore, J.R. (1994): Antioxidant status of dairy cows supplemented prepartum with vitaminE and selenium, Journal of Animal Science 77:3087-3095
- Buettner, G.R. (1993): The pecking order of free radicals and antioxidants: lipid peroxidation, alpha tocopherol, and ascorbate, Archives of Biochemistry and Biophysics 300, 535-543
- Calamari, L., Maianti, M.G., Amendola, F., Lombardi (1999): On some aspects of the oxidative status and on antioxidants in blood of dairy cows during summer, Pages 449-451 in Proc. 13th Associazione Scientifica Produzioni Animali Congress, Piacenza, Italy
- Cao, G., Prior, R.L. (1998): Comparison of different analytical methods for assesing total antioxidant capacity of human serum, Clinical Chemistry 44,1309-1315
- Castillo, C., Benedito, J.L., Lopez-Alonso, M., Miranda, M., Hernandez, J. (2001): Importancia del estres oxidativo en ganado vacuno:en relacion con el estado fiziologico (prenez y parto) y la nutricion, Archivos de Medicina Veterinaria 33, 5-20
- Castillo, C., Hernandez, J., Bravo, A., Lopez-Alonso, M., Pereira, V., Benedito, J.L. (2005): Oxidative status during late pregnancy and early lactation in dairy cows, The Veterinary Journal 169, 286-292
- Castillo, C., Hernandez, J., Lopez-Alonso, M., Miranda, M., Benedito, J.L. (2003): Values of plasma hydroperoxides and total antioxidant status in healthy dairy cows: preliminary observations, Archives of Animal Breeding 46, 227-233
- Castillo, C., Hernandez, J., Valverde, I., Pereira, V., Sotillo, J., Lopez Alonso, M., Benedito, J.L: (2006): Plasma malonaldehyde (MDA) and total antioxidant status (TAS) during lactation in dairy cows, Research in Veterinary Science 80, 133-139
- Chen, K., Suh, J., Carr, A.C., Morrow, J.D., Zeind, J.,Frei, B. (2000): Vitamin C supresses oxidative lipid damage in vivo, even in the presence of iron overload, American Journal of Physiology, Endocrinology and Metabolism 279, E1406-E1412
- Chevion, M., Berenshtein, E., Stadtman, E.R. (2000): Human studies related to protein oxidation: protein carbonyl content as a marker of damage, Free Radical Research 33, S99-S108
- Chiradia, E., Avellini, L., Rueca, F., Spaterna, A., Porciello, F., Antonioni, M.T., Gaiti, A. (1998): Physical exercise oxidative stress and muscle damage in racehorses, Comp. Bioch. and Physiol. B 119, 833-836
- Clemens, M.C., Waller, H.D. (1987): Lipid peroxidation in erythrocytes, Chem. Phys. Lipids. 45:251-268
- Collins, A.R. (2004): The comet assay for DNA damage and repair: principles, applications, and limitations, Molecular Biotechnology 26, 249-261
- Curtis, M.A. (1997): Epidemiology of uterine infection in dairy cows: antioxidant and metabolic investigations, PhD Thesis, University of Sydney, Australia
- DiGuiseppi, J., Fridovich, I. (1984): The toxicity of molecular oxygene, Crit. Rev. Toxicol. 12:315-342
- Doepel, L., Lapierre, H., Kenelly, J.J. (2002): Peripartum performance and metabolism in dairy cows in response to prepartum energy and protein intake, J. Dairy Sci. 85, 2315-2334
- Drackley, J.K. (1999): Biology of dairy cows during the transition period:The final frontier? J. Dairy Sci. 82:2259-2273
- Esterbauer, H., Zollner, H. (1989): Methods for determination of aldehydic lipid peroxidation products, Free Radical Biology and Medicine 7, 197-203
- Frei, B., Stocker, R., Ames, B.N. (1998): Antioxidant defenses and lipid peroxidation in human blood plasma, Proc. Natl. Acad. Sci. USA 85:9748-9752
- Ghiselli, A., Serafini, M., Natella, F., Scaccini, C. (2000): Total antioxidant capacity as a tool to asess redox status: critical view and experimental data, Free Radical Biologicla Medicine 29, 1106-1114
- Goff, J.P., Horst, R.L. (1997): Physiological changes at parturition and their relationship to metabolic disorders, J. Dairy Sci. 80:1260-1268
- Gröhn, Y.T., Erb, H.N., McCulloch, C.E., Saloniemi, H.S. (1989): Epidemiology of metabolic disorders of dairy cattle: Association among host characteristics, disease, and production, J. Dairy Sci. 72:1876-1885
- Grum, D.E., Drackley, J.K., Hansen, L.R., Cremin, Jr. J.D. (1996): Production, digestion and hepatic lipid metabolism of dairy cows fed increased energy from fat or concentrate, J. Dairy Sci. 79, 1836-1849
- Grummer, R.R. (1993): Etiology of lipid-related metabolic disorders in periparturient dairy cows, J. Dairy Sci. 76:3882-3896
- Halliwell, B., Gutteridge, J.M.C. (1999): Free Radicals in Biology and Medicine, third ed. Oxford University Press, New York
- Harmon, R.J., Lu, M., Trammel, D.S., Smith, B.A. (1997): Influence of heat stress and calving on antioxidant activity in bovine blood, J. Dairy Sci. 80(Suppl. 1):264 (Abstr.)
- Harrison, J.P., Hancock, D.D., Conrad, H.R. (1984): Vitamin E and selenium for reproduction in dairy cows, J. Dairy Sci. 67:123-131
- Hermes-Lima, M., Storey, J.M., Storey, K.B. (1998): Antioxidants defenses and metabolic depression. The hypothesis of preparation for oxidative stress in land snails, Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 120:437-448
- Ingvartsen, K.L., Andersen, J.B. (2000): Integration of metabolism and intake regulation: a revew focusing on peripartureint animals, J. Dairy Sci. 83, 1573-1597
- Kankofer, M. (2001): The levels of lipid peroxidation products in bovine retained and non retained placenta, Prostaglandins, Leukotriens and Essential Fatty Acids 64, 33-36
- Ketelaars, J.J.M.H., Tolkamp, B.J. (1996): Oxygen efficiency and the control of energy flow in animals and humans, Journal of Animal Science 74, 3036-3051
- Kowaliszyn B., Sitkowska Beata, Mroczkowski S. (2008): Physiological state of dairy cattle and somatic cells count class versus level of plasma malonyldialdehyde, Journal of Central European Agriculture, Vol. 9, No. 2, 373-376
- Kusmic, C., Picano, E., Buscetti, C.L., Petersen, C., Barsacchi, R. (2000): The antioxidant drug dipyridamole spares the vitamin E and thiols in red blood cells after oxidative stress, Cardiov. Res. 47:510-514
- Lomba. F. (1996): Influence of dietary cation-anion and oxidative-antioxidans balances on diseases occuring around parturition in the dairy cows, Ann. Med. Vet. 140:109-122
- Lykkesfeldt, J. (2001): Determination of malondialdehyde dithiobarbituric acid adduct in biological samples by HPLC with fluorescence detection: Comparison with UV-visible spectrophotomoetry, Clinical Chemistry 47, 1725-1728
- Lykkesfeldt, J. (2002); Measurement of ascorbic acid and dehydroascorbic acid in biological samples, In: Maines, M., Costa, L.G., Hodson, E., Reed, D.J., Sipes, I.G. (Eds.), Current Protocols in Toxicology, John Wiley and Sons, New York, pp. 7.6.1.-7.6.1.5.
- Lykkesfeldt, J., Svendsen, O. (2007): Oxidants and antioxidants in disease: Oxidative stress in farm animals, The Veterinary Journal 173, 502-511
- Lykkesfeldt, J., Viscovich, M., Poulsen, H.E. (2003): Ascorbic acid recycling in human eritrhorcytes is induced by smoking in vivo, Free Radical Biology and Medicine 35, 1439-1447
- Marcos, E., Mazur, A., Cardot, P., Rayssiguier, Y. (1990): The effect of pregnancy and lactation in serum lipid and apolipoprotein B and A-I levels in dairy cows, Journal of Animal Physiology and Animal Nutrition 64, 133-138
- Miller, J.K., Brzezinska-Slebodzinska, E., Madsen, F.C. (1993): Oxidative stress, antioxidants and animal function, J. Dairy Sci 76: 2812-2823
- Miller, J.K., Madsen, F.C., Lyons, T.P., Jacques, K.A. (1994): Transition metals, oxidative status, and animal health: Do alterations in plasma fast-acting antioxidant lead to disease in livestock? Biotechnology in the feed industry. Pages 283-301 in Proc. Alltech's Tenth Annu. Symp. T.P. Lyons and K.A. Jacques, ed. Nottingham Univ. Press, Nottingham, UK
- Minor, d.J., Trower , S.L., Strang, B.D., Shaver, R.D., Grummer, R.R. (1998). Effects of nonfiber carbohydrate and niacin on periparturient metabolic status and lactation in dairy cows, J. Dairy Sci. 81, 189-200
- Morrow, J.D. (2000): The isoprostanes: Their quantification as an index of oxidant stress status in vivo, Drug Metabolism Reviews 32, 377-385
- Nardone, A., Lacetera, G., Bernabucci, U., Ronchi, B. (1997): Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and early postpartum period, J. Dairy Sci. 80:838-844
- Nikki, E., Noguchi, N. (2000): Evaluation of antioxidant capacity is being measured by which method? IUBMB Life 50, 323-329
- Noble R.C., O'Kelly, J.C., Moore, J.H. (1976): Observations on changes in lipid composition and lecitin-cholesterol-acyl transferase reaction of bovine plasma induced by heat exposure, Lipids 8:216-223
- NRC - Nutritional Research Council (1989): Nutrient Requirements of Dairy Cattle, sixth revised edition, National Academic Science, Washington, DC
- Park, A.F., Shirley, J.E., Titgemeyer, E.C., Meyer, E.J., VanBaale, M.J., Vanderhaar, M.J. (2002): Effect of protein level in prepartum diets on metabolism and performance indairy cows, J. Dairy Sci. 85, 1815-1828
- Payne, C.M., Bernstein, C., Bernstein, H. (1995): Apoptosis overview emphasizing the role of oxidative stress, DNA dmaage and signal-transduction pathways, Leukemia and Lymphoma 19, 43-93
- Poulsen, H.E. (2005):Oxidative DNA modifications, Experimental and Toxicologic Pathology 57 (Suppl. 1), 161-169
- Rosenkranz, A.R., Schmadienst, S., Stuhlmeier, K.M. Chen, W., Knapp, W., Zlabinger, G.J. (1992): A microplate assay for the detection of oxidative products using 2',7'-dichlorofluorescin-diacetate, Journal of Immunological Methods 156, 39-45
- Roth, R. (2000): Oxygen free radicals and their clinical implications, Acta Chirurgica Hungarica 36, 302-305
- Sies, H. (1991): Oxidative stress, Academic Press Ltd., Orlando, FL.
- Smith, K. L., Harrison, J.H., Hancock, D.D., Todhunter, D.A., Conrad, H.R. (1984): Effect of vitamin E and selenium supplementation on incidence of clinical mastitis and duration of clinical symptoms, J. Dairy Sci. 67:1293-1299
- Šamanc, H., Stojić, V., Kirovski, Danijela, Jovanović, M., Cernescu, H., Vujanac, I. (2010): Thyroid Hormones Concentration during the Mid-Dry Period: An Early Indicator of Fatty Liver in Holstein-Friesian Dairy Cows, Journal of Thyroid Research, Vol. 1, 2010 (in press)
- Tokoyuni, S., (1999): Reactive oxygen species-induced molecular damage and its application in pathology, Pathol. Int. 49:91-102
- Trout, J. P., McDowell, L.R., Hansen, P.J. (1998): Characteristics of the estrous cycle and antioxidant status of lactating Holstein cows exposed to heat stress, J. Dairy Sci. 81:1244-1250
- Turk, R., Juretic, D., Geres, D., Turk, N., Rekic, B., Simeon-Rudolf, V., Svetina, A. (2004): Serum paraoxonase activity and lipid parameters in the early postpartum period of dairy cows, Research in Veterinary Science 76, 57-61
- Vanderhaar, M.J., Yousif, G., Sharma, B.K., Herdt, T. H., Emery, R.S., Allen, M.S., Liseman, J.S. (1999): Effect of energy and protein density of prepartum diets on fat and protein metabolism of dairy cattle in the peripartum period, J. Dairy Sci. 82:1282-1295
- Vernon, R.G. (2002): Nutrient partitioning, lipid metabolism and relevant imbalances. In: Kaske, M., Scholz, H., Holtershinken, M. (Eds.), Recent Developments and Perspectives in Bovine Medicine, Klinik fur Rinderkrankheiten, Tierartzliche Hochschule, Hannover, pp 210-223
- Watcher, C.M., McDaniel, B.T., Whitlow, L.W., Pettyjohn, S. (1999): Genetics of antioxidant activity in Holsteins and Jerseys: association with various traits, J. Dairy Sci. 82 (Suppl. 1), 31.
- Webster, A.J.F. (1991): Metabolic responses of farm animals to high temperature, EAAP Publ., 55:15-22
- Xu, S., Harrison, J.H., Chalupa, W., Sniffen, C., Julien, W., Sato, H., Fujieda, T., Watanabe, K., Udea, T., Suzuki, H. (1998): The effect of ruminal bypass lysine and methionine on milk yielld and composition of lactating cows, J. Dairy Sci. 81, 1062-1077
preuzmi
seminarski rad u wordu » » »
