ДРУГИ СЕМИНАРСКИ РАБОТИ
- МЕДИЦИНА: |
||||||||||||||||||||
|
||||||||||||||||||||
Клонирање на ДНА
Во последните години (од клонирањето на овцата Доли па навака) зборот клон е еден меѓу најкористените во секојдневниот речник. Меѓутоа голем дел од оние кои го користат зборот клон или зборот клонирање, или не го знаат или само го погодуваат неговото вистинско значење.
Во овој прилог ќе ги наведеме поважните негови значења и ќе одговориме на едни од најпоставуваните прашања.
Што е клон?
Збир од генетски еднакви организми, создаден од еден родител по пат на неполово размножување. Пример за вакво размножување е пупењето кај растенијата.Што e клонирање?
По дефиниција клонирањето претставува процес на формирање популација на клетки со идентичен генотип. Молекуларното клонирање во потесна смисла на зборот се однесува на in vivo амплификација со која се продуцираат енормен број идентични молекули.Како se врши клонирањето? Преку рекомбинација, односно инсерција на егзогена (туѓа) DNA во автономно реплицирачки вектор. Рекомбинираниот вектор со DNA инсертот се користи за трансформација на "домаќини" какви што се: бактериите, инсектите, мамалиските или други клeтки.DNA сегментот се реплицира заедно со векторот и како резултат на тоа се добиваат голем број идентични копии - клонови.
Со чија помош се врши клонирањето? Во текот на овие процедури, како молекуларни алатки се користат повеќе ензими какви што се:
• Рестрикциските ендонуклеази за пресекување на двоверижната DNA,
• Лигазите за нејзино лепење преку повторно воспоставување на фосфодиестерски врски,
• Фосфатази за превенција на самолигирање на векторот и др. Планетарно познат резултат од примената на оваа техника е овцата Доли.
Сл.1.1. Постапка при клонирање
Рвкомбинирачка DNA швхнологијаСакаме да подготвиме субклонирани DNA примероци т.е. пораснати мултипни копи на определено парче од DNA за понатамошни субклонирања или секвенционирања. Приоритети во тој случај би ни биле субклонирачките процедури по редослед на важноста.
1. да бидеме во можност да ги препознаеме сите региони во геномот
2. да продуцираме примероци од секвенционирана DNA
3. Ако како резултат би сакале да завршиме со добивање на ГеномскатаБиблиотека, исто така ќе треба да ги подредиме клоновите низ геномот т.е. да ги детерминираме релативните позиции на клоновите.
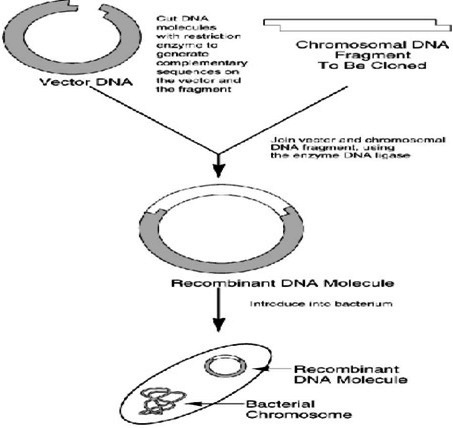
Сега да претпоставиме дека сме запознаени со даден геном и дека сме тргнати по неговата основна композиција. Неговата DNA исто така е претставена како туѓа или донирачка DNA. Идејата е да направиме рекомбинантна DNA со пресекување на донирачката DNA, инсертирајќи даден фрагмент во мал реплицирачки молекул (вектор) под одредени услови. Потоа ќе го засилиме фрагментот по неговата должина, што ќе резултира со молекуларен клон од внесената DNA молекула. Векторските молекули со нивните инсертирани делови се наречени рекомбинантни DNA молекули бидејќи тие претставуваат комбинации од донорска DNA со DNA вектори од комплетно различен ресурс (најчесто бактериски пласмид или вирус) рекомбинираната DNA структура потоа се користи за да ги трансформира бактериските клетки и нивните заеднички својства во единствен рекомбинирачки вектор. Бактериските клетки потоа се засадуваат и им се дозволува да растат во колонии и од една индивидуална трансформирана клетка со единствен рекомбиниран вектор ќе пораснат колонии со милиони клетки кои ќе го содржат истиот рекомбиниран вектор. Затоа една индивидуална колонија претставува огромна популација на идентични DNA делови и оваа популација е наречена ДНА клон.
Процвдури ири клонирањв на DNA
1) Изолирање на DNA:
Првиот чекор е да изолираме донор и вектор. Методот за изолирање на геномска DNA (ова е типот на DNA кој треба да го добиеме од донорот) кој постоел долго пред изумот со рекомбинираната DNA технологија. Процедурите за добивање на DNA вектор зависи од самата природа на векторот. Бактериските плазмиди се често користени и претходно мора да бидат исчистени од геномската бактериска DNA. Еден од можните протоколи е базиран на набљудувањето дека специфична алкална рН ја денатурира бактериската геномска DNA, но не ги денатурира плазмидите.2) Сечење на DNA:
Откривањето на карактерот на рестриктивните ензими ја направи техниката на рекомбинирање возможна. рестриктивните ензими се продуцирани како резултат на одбрамбените механизми кои бактериите ги развиваат против фагите. Тие го претставуваат бактерискиот имун систем. Ензимите пак ги инактивираат фагите со сечење на нивната DNA на местата за ресртикција. Местата за рестрикција се специфижни целни секвенци кои се полиндромски (имаат иста нуклеотидна секвенца но во антипаралелен правец) и ова е само една од многуте особини кои ги прават соодветни за DNA манипулација. ПРИМЕР:
Ензимот за рестрикција EcoRY (E.Coli) ги препознава следните 6 нуклеотидни пар секвенци во DNA на било која организам:
Ензимот ги сече секвенците во правец помеѓу G и A нуклеотидите:
Сето ова не би било можно без т.н.р. "лепливи" делови кои имаа способност да ја хидрогенизираат, залепат комплементарната секвенца. Сега ако два различни DNA молекули се пресечат со истиот рестриктивен ензим, двете ќе произведат фрагменти со комплементарни лепливи краеви, со што ќе овозможат почеток на DNA рекомбинацијата.3) Лепење на DNA:
Донорот DNA и векторот DNA сега се пресечени со истиот рестриктивен ензим и измешани во тест сад со цел да се овозможи да се залепат "лепливите" краеви и да ја формираат рекомбинираната DNA. Во оваа фаза гликоза фосфатот не е комплетиран во двете позиции во секој спој. Како идае фрагментите можат да бидат поврзани засекогаш со додавање на ензимот DNA лигаза, која креира диестер на фосфор на споените краеви за да ја направи DNA молекулата продолжена. Еден од проблемите кој може да настане во овој дел од процесот на клонирање на DNA е тоа што пресечените краеви на молекулот можат да се здружат повторно а да не формираат рекомбинирана DNA. Со цел да се реши проблемот се додава ензимот завршна трансфераза, која го кактализира додавањето на нуклеотидни опашки на 3' краевите од DNA синџирот.
4) Зајакнување на рекомбинираната DNA:
Рекомбинираната плазмидска DNA е воведена во клетката домаќин со помош на трансформација. Во клетката домаќин векторот ќе се реплицира нормално, сега донор DNA - та е автоматски реплицирана заедно со векторот. Секој рекомбиниран пласмид кој влегува во состав на клетката ќе формира мултипни копи на самиот себе. Субсеквенционално со многуте циклуси на делба клетката ќе се зголеми и рекомбинираниот вектор ќе направи повеќе репликации. Бактериската колонија на крај ќе содржи милијарди копии на самостојни долови од донорската DNA. Овој сет на зајкнати копи на самостојни донорски DNA молекули е всушност DNA клон.
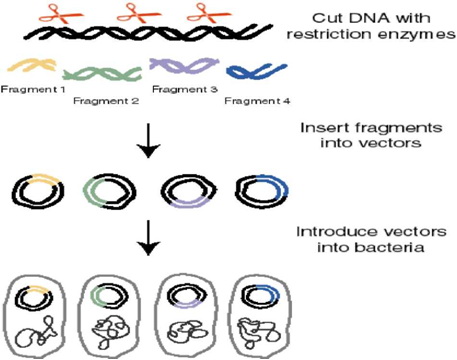
Сл.1.3. Клонирање на DNA
ДНЛ библиошвки
Различни извори на DNA можат да бидат искористени за да создадеме рекомбинантни DNA молекули.
Колекцијата од рекомбинантни DNA молекули создадени од специфичен вектор се нарекува ДНЛ библиотвка. DNA библиотеката од човечкиот геном користи плазмиди како вектори кои и се потребни за одржи неколку стотици илјади клонови. Библиотеките овозможуваат подобро познавање на еволуцијата, експресијата и регулацијата на хуманата DNA, тие овозможуваат детална анализа на DNA на одредени хромозоми, локализација и број на хромозоми, а со тоа и запознавање на генетската мапа на хромозомите.
Избор на ввкшор и на организам -" домаќин" za молвкуларно клонирањвИзборот на комбинацијата на вектор и на клетка - домаќин за клонирање на DNA секвенца зависи од повеќе фактори. Векторите теба да бидат релативно мали макромолекули за да можат едноставно да се манупулираат и да можат да се реплицираат во домаќинските клетки.
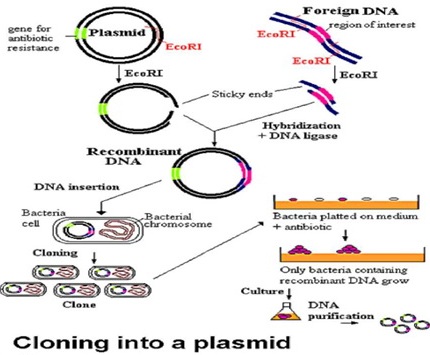
Постојат големи разлики и во однос на едноставноста и во однос на економичноста на методите за пропагирање на "домаќинските" клетки во култура in vivo, при што бактериите и габите се многу полесни и поевтини за манипулација во лабораториски услови, отколку вишите еукариотски клетки. Од тие причини, изборот на комбинација на вектор и на домаќин најчесто е компромис меѓу нивните карактеристики и ограничувања. Во процесите на молекуларно клонирање најчесто се користат следниве вектори:• Плазмиди: бактериските плазмиди се релативно мали, циркуларни ДНА молекули кои егзистираат екстрахромозомално од 10, па се до 700 копии во бактериските клетки и се реплицираат под "релаксирана" контрола т.е. релативно независно од репликацијата на бектерискиот геном. При клонирање на DNA, DNA фрагментот кој го содржи генот за кој сме заинтересирани, го внесуваме во клонирачкиот вектор или пласмид. Оние нишки од генот кои сакаме да ги издвоиме, ги сечеме со рестрикциска ендонуклеаза. Тогаш плазмидот се отвара и гените се ослободуваат од родителската DNA нишка (DNA молекулот се линеаризира). Плазмидите исто така имаат и комплементарни "лепливи" (кохезивни) краеви. Инсерцијата на фрагментите во плазмидот е возможна благодарение на компатибилноста на кохезивните краеви добиени со дигестија на идентичниот ензим. Отворениот пласмид и ослободениот ген се мешаат со DNA лигаза при што се добиваат рекомбинантни вектори кои се користат за трансформација на соодветни бактериски клетки кои се размоножуваат со инкубација во соодветни услови. При тоа се умножуваат и рекомбинираните плазмиди, а со нив и инсертираната DNA се мултиплицира во милијарди идентични копи -клонови. Покрај мултиплицирањето на DNA инсертите, со клонирањето може да се врши и експресија на клонираниот ген и биотехнолошки да се произведуваат индустриски количества на важни протеини, каков што е хуманиот инсулин, на пример.
Сл. 1.4. Клонирање во плазмиди
• Вирусни вектори: вирусите имаат определени предности како вектори за клонирање на DNA. Тие ефикасно ги инфицираат приемчивите клетки и поради тоа можат да ја инсертираат туѓата DNA со многу поголема фрекфенција отколку плазмидите. Бакуловирусот на пример инфицира клетки од инсекти, па често се користи за клонирање и експресија на еукариотски клетки во културата на некои инсектни клетки. Ретровирусите се користат за стабилана интеграција во геномот на клетки од цицачи и континуирана експресија на клонираниот ген. определени генски модифицирани ретровируси и аденовируси се применуваат и за т.н. генска терапија, односно за корекција на мутиран или за внесување на соодветен ген при некое заболување.• Космиди: векторите кои се хибриди на ламбда фагот(бактериски вирус кој често се користи како вектор за генско клонирање) и на плазмидите се означуваат како космиди. Тие можат да се реплицираат автономно како и плазмидите, а можат да се пакираат постабилно и компактно како и ламда фагот. Покрај тоа можат и да пренесуваат околу три пати поголеми DNA инсерти отколку ламбда фагот, односно од 30 до 45 kb. Фрагемидите се композитни вектори добиени со спојување на едноверижни фаги со некои плазмиди. Во зависност од условите за раст, фрагемидите се реплицираат како едноверижни или двоверижни DNA молекули, што е важно при определени методи за клонирање.
• Вештачки хромозоми: тоа се синтетски конструирани вектори кои содржат DNA секвенци неопходни за формирање на структура на хромозом: теломере, центромера, регион за почеток на репликација и други организациски елементи. Се користи во геномиката за инсерција на енормно големи секвенци на DNA. Кон нив се однесуваат бактерискиот вештачки хромозом - BAC ( Bacterial Arteficial Chromosome) , габичниот вештачки хромозом - FAC (Fungal Arteficial Chromosome), квасниот вештачки хромозом - YAC (Yeast Arteficial Chromosome) и други вектори.
Рекомбинираните вектори со инсертирана DNA се користат за трансформација на
следниве клетки-домаќини:
• Прокариоти (бактерии): бактериите се традиционално први и најчесто користени домаќини за трансформација со рекомбинирани вектори. Големи предности во користењето на бактерии за тие цели се релативно едноставното и евтино култивирање, како и способноста за раст во голема биомаса, а со тоа и ефикасно умножување на инсертираниот ген или генски сегмент. Освен тоа, механизмите на регулација на генската експресија се доста добро проучени и подложени на контрола. Од друга страна, бактериите не се добар избор како домаќини кај експериментите при кои се предвидува експресија на клониран еукариотски ген. прокариотите не се способни за посттранскрипциски модификации како што е DNA сплајсингот, па присуството на интрони го оневозможува правилното процесирање на mRNA молекули.
• Еукариоти: во техниките на рекомбинантна ДНА се користат повеќе типа еукариотски клетки. Нижите еукариоти, какви што се квасните габи, значително се разликуваат по своите карактеристики од вишите еукариотски клетки, па затоа имаат и различни индикации за примена како домаќини за трансформација со рекомбинирани вектори при молекуларно клонирање.
Квасните габи (како што е Sachromyces cerevisiae) се култивираат мошне слично како и бактериите т.е. релативно економично, едноставно и ефикасно. Механизмите на регулација на на генската експресија се солидно познати и доста лесно се контролираат in vitro. Габите имаат механизми за отстранување на интроните од RNA транскриптите, но не се секогаш компатибилни со организацијата на посложените гени од вишите еукариоти. Ограничување е и тоа што габите не вршат посттранслациски модификации како што е гликозилацијата, на еднаков начин како што го прават тоа мамилските клетки. Но, еукариотските, особено мамилските клетки се многу покомплицирани и поскапи за култивирање, а и тешко разраснуваат во голем број. Поради сеуште недоволно проучената регулација на генската експресија, комплицирана е манипулацијата со ниваната експресија на клонираниот ген.
Заклучок
Важнос на DNA клонирање
Во историјата на медицинската генетика клонирањето на DNA е едно од најзначајните откритија во молекуларната биологија и е незамнелива процедура во генетскиот инженеринг и DNA технологијата. "Пробивањето" на хромозомите во средината на 1950 година беше револуционерно. Пред DNA клонирањето нашето знаење за DNA беше екстремно ограничено.
Технологијата на DNA клонирањето влијаеше на измените на сето претходно знаење, и ја револуционираше генетиката како наука. За да разбереме зошто е тоа така треба да се свати огромната големина и сложеноста на DNA секвенците (споредени на пример со протеински молекули). Во последните две декади DNA технологијата имаше многу голем ефект не само во медицинската генетика, туку и во сите подрачја на биолошката наука.
Идентификацијата на рестриктивната ендонуклеаза, развојот на клонирачките DNA вектори, појавата на Southern blot техника во 1970 година и многу други скорешни достигнувања на полимеразната верижна реакција, се рангираа како основни во развојот на ова поле.
Со техниките на генетскиот инженеринг, протеините кои се експресираат можат да се модифицираат или да се дизајнираат преку генско манипулирање со DNA секвенцата која го кодира протеинот од интерес. Што е од многу голем интерес како за медицината така и за човештвото воопшто затоа што со примената на оваа техника - ДНА клонирањето можеме да ги одвоиме оние протеини за кои сме заинтересирани, на пример протеини кои ни се потребни за испитување на резистентноста кон некои антибиотици или протеинот за синтеза на инсулин како и многу други.
Користена литература:
1. Caшо Панов. Основни методи во молекуларната биологија.
Универзитет "Св. Кирил и Методиј". Природно Математички Факултет - Скопје, 2003.
2. Robert F. Mueller, Ian D. Young. Emery's Elements of Medical Genetics. Eleventh edition, Churchill Livingstone, New York 2001.
3. Tom Strachan & Andrew P. Read. Human Molecular Genetics 3. Garland Science, London and New York, 2004.
4. Владимир E. Трајковски. Хумана Генетика. Универзитет Св. "Кирил и Методиј" - Скопје, 2005
5. D-r med. Ivan Skeling. Medicina HR www. Medicina.hr/clanci/klon.html
6. Simon Cawley . Genome Projects www.berkeley.edu/users/terry/Classes.html
7. National health museum. Cloning DNA www.accessexcellence.org/AB/GG/plasmid.htlm
Download СЕМИНАРСКА РАБОТА у wordu » » »

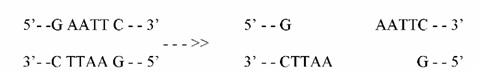
Besplatni Seminarski Radovi - Бесплатно семинарска работа